![]() プラズマクラスター技術
プラズマクラスター技術
 ニュース一覧
ニュース一覧
ドイツ アーヘン応用科学大学 アートマン教授との共同研究
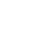
プラズマクラスターイオン ※1による細菌不活化メカニズムを解明
シャープは4年前の2000年にプラスとマイナスのイオンを放出して空中除菌するプラズマクラスターイオン技術を開発し、世界の学術研究機関と共同で、イオンが浮遊する細菌・ウイルス・アレルゲンなどの水素を抜きとって不活化すること、およびその減少率を数値的に検証してきました。
今回は単に細菌やウイルスに対する効能検証の域を超えてこのアカデミックマーケティング※2をさらに深化させ、細胞工学分野の世界的権威であるドイツ アーヘン応用科学大学 細胞生物物理・生物工学部 ゲハート アートマン教授(Prof. Gerhard Artmann)と2002年から共同研究を実施し、プラズマクラスターイオンが細菌の表面細胞膜のタンパク質を破壊し不活化するメカニズムを解明しました。
細胞質に含まれるタンパク質は生命活動を行う必須物質です。一般に細菌は表面細胞膜の約70%がタンパク質であり、ウイルスは中心の遺伝子をタンパク質が取り囲んだ構造であることが知られています。今回の研究結果によって表面にタンパク質をもつ広範囲の浮遊有害菌などへの効果が期待できることが学術的に解明されました。
アートマン教授の研究グループと共同で、不活化された細菌のタンパク質を分子生物学の手法で解析し併せて電子顕微鏡で観察した結果、細菌はプラズマクラスターイオンにより60種類以上もの表面細胞膜のタンパク質を破壊され、表面細胞膜が機能不全となり不活化することが証明されました。
また紫外線や放射能など生体内部に侵入して殺菌するものとは異なり、プラズマクラスターイオンは細菌内部のDNA(遺伝子)を損傷しない(細胞がガン化しない)ことも確認できております。
こうしたことから、プラズマクラスターイオン技術が、効能の広さと安全性において他の空中除菌方式を凌駕する優位性が分子生物学の分野からも証明されました。そして空気浄化技術において細菌不活化メカニズムを解明したのは、家電業界でシャープだけです。
- ※1 プラズマクラスター、プラズマクラスターイオンおよびPlasmaclusterはシャープ株式会社の商標です。
- ※2 技術の効能について、先端の学術研究機関と共同で科学的データを検証し、それをもとに商品化を進めるマーケティング手法。
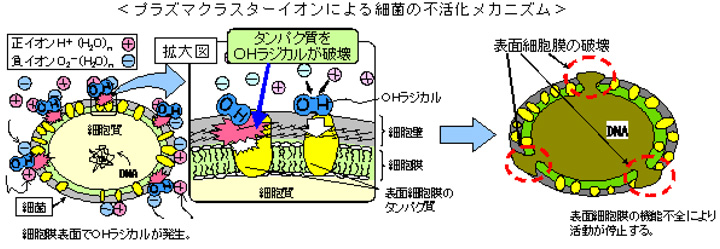
細菌の不活化写真
容積4Lのボックス内にプラズマクラスターイオン発生素子を設置し、細菌に一定時間イオンを照射後、細菌を電子顕微鏡で分析しました。その結果、イオン照射により細胞膜が破れ、内部が出てきていることが確認されました。これはイオンが細菌の表面細胞膜のタンパク質を切断し、膜に穴が開いたことを意味します。これより細菌の不活化は膜機能不全によるものと確認され、不活化した細菌は活動を停止し増殖しません。
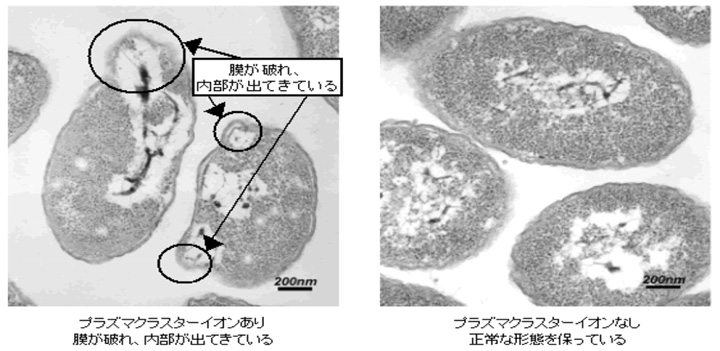
細菌の表面細胞膜のタンパク質の分析
容積4Lのボックス内にプラズマクラスターイオン発生素子を設置し、細菌に一定時間イオンを照射後、細菌の表面細胞膜のタンパク質を2次元電気泳動法※3で分析しました。イオン照射していない表面細胞膜では多数のタンパク質が存在していますがイオン照射により表面細胞膜のタンパク質がほとんどなくなり、60種類以上ものタンパク質のシグナルが消失することを確認しました。これはイオンが細菌の表面細胞膜のタンパク質を破壊したことを意味します。
アカデミックマーケティングによるプラズマクラスターイオン効能一覧
| 対象有害物質 | 種 類 | 実 証 機 関 | 時 期 |
|---|---|---|---|
| 真菌 | クラドスポリウム | (財) 石川県予防医学協会 | 2000年 9月 |
| ドイツ リューベック医科大学(増殖抑制効果) | 2002年 2月 | ||
| CT&T(ドイツ アーヘン応用科学大学 アートマン教授) | 2004年11月 | ||
| ペニシリウム、アスペルギルス | ドイツ リューベック医科大学(増殖抑制効果) | 2002年 2月 | |
| アスペルギルス、ペニシリウム(2種)、 スタキボトリス、アルテルナリア、ムーコル |
CT&T(ドイツ アーヘン応用科学大学 アートマン教授) | 2004年11月 | |
| 細菌 | 大腸菌 | (財) 石川県予防医学協会 | 2000年 9月 |
| 大腸菌、ブドウ球菌、カンジダ菌 | 中 国 上海市予防医学研究院 | 2001年10月 | |
| バチルス菌 | (財) 北里環境科学センター | 2002年 9月 | |
| CT&T(ドイツ アーヘン応用科学大学 アートマン教授) | 2004年11月 | ||
| MRSA (メチシリン耐性黄色ブドウ球菌) |
(財) 北里環境科学センター | 2002年 9月 | |
| (社) 北里研究所 北里研究所メディカルセンター病院 | 2004年 2月 | ||
| シュードモナス、エンテロコッカス、スタフィロコッカス | ドイツ リューベック医科大学 | 2002年 2月 | |
| エンテロコッカス、スタフィロコッカス、サルキナ、マイクロコッカス | CT&T(ドイツ アーヘン応用科学大学 アートマン教授) | 2004年11月 | |
| アレルゲン | ダニ、花粉 | 広島大学大学院 先端物質科学研究科 | 2003年 9月 |
| 浮遊アレルゲン | カナダ喘息協会 | 2004年 4月 | |
| ウイルス | インフルエンザウイルス | (財) 北里環境科学センター | 2002年 9月 |
| 韓 国 ソウル大学 | 2003年 9月 | ||
| 中 国 上海市予防医学研究院 | 2003年12月 | ||
| (社) 北里研究所 北里研究所メディカルセンター病院 | 2004年 2月 | ||
| コクサッキーウイルス | (財) 北里環境科学センター | 2002年 9月 | |
| ポリオウイルス | (財) 北里環境科学センター | 2002年 9月 | |
| コロナウイルス | (社) 北里研究所 北里研究所メディカルセンター病院 | 2004年 7月 |
| 対象有害物質 | 種類 | 実証機関 | 時期 |
|---|---|---|---|
| 真菌 | クラドスポリウム | (財) 石川県予防医学協会 | 2000年 9月 |
| ドイツ リューベック医科大学(増殖抑制効果) | 2002年 2月 | ||
| CT&T(ドイツ アーヘン応用科学大学 アートマン教授) | 2004年11月 | ||
| ペニシリウム、アスペルギルス | ドイツ リューベック医科大学(増殖抑制効果) | 2002年 2月 | |
| アスペルギルス、ペニシリウム(2種)、 スタキボトリス、アルテルナリア、ムーコル |
CT&T(ドイツ アーヘン応用科学大学 アートマン教授) | 2004年11月 | |
| 細菌 | 大腸菌 | (財) 石川県予防医学協会 | 2000年 9月 |
| 大腸菌、ブドウ球菌、カンジダ菌 | 中 国 上海市予防医学研究院 | 2001年10月 | |
| バチルス菌 | (財) 北里環境科学センター | 2002年 9月 | |
| CT&T(ドイツ アーヘン応用科学大学 アートマン教授) | 2004年11月 | ||
| MRSA (メチシリン耐性黄色ブドウ球菌) |
(財) 北里環境科学センター | 2002年 9月 | |
| (社) 北里研究所 北里研究所メディカルセンター病院 | 2004年 2月 | ||
| シュードモナス、エンテロコッカス、スタフィロコッカス | ドイツ リューベック医科大学 | 2002年 2月 | |
| エンテロコッカス、スタフィロコッカス、サルキナ、マイクロコッカス | CT&T(ドイツ アーヘン応用科学大学 アートマン教授) | 2004年11月 | |
| アレルゲン | ダニ、花粉 | 広島大学大学院 先端物質科学研究科 | 2003年 9月 |
| 浮遊アレルゲン | カナダ喘息協会 | 2004年 4月 | |
| ウイルス | インフルエンザウイルス | (財) 北里環境科学センター | 2002年 9月 |
| 韓 国 ソウル大学 | 2003年 9月 | ||
| 中 国 上海市予防医学研究院 | 2003年12月 | ||
| (社) 北里研究所 北里研究所メディカルセンター病院 | 2004年 2月 | ||
| コクサッキーウイルス | (財) 北里環境科学センター | 2002年 9月 | |
| ポリオウイルス | (財) 北里環境科学センター | 2002年 9月 | |
| コロナウイルス | (社) 北里研究所 北里研究所メディカルセンター病院 | 2004年 7月 |
ゲハート アートマン教授(Prof. Gerhard Artmann)のプロフィール

ドイツ アーヘン応用科学大学 細胞生物物理・生物工学部 教授
ドイツ アーヘン応用科学大学 応用物理学部 教授
米 国 ドレクセル大学 助教授
CT&T(細胞組織技術研究所) 代表
理学博士
[専門]
細胞工学(分子レベルで生物を研究)、生物物理学
[経歴]
1993年~1994年 中国 上海医学大学生物物理学部客員教授 1994年~1997年 米国 カリフォルニア大学サンディエゴ校客員研究員
1998年 中国 重慶大学顧問教授
2000年~2001年 米国 カリフォルニア大学サンディエゴ校客員研究員
[活動]
1991年~ ドイツ アーヘン応用科学大学細胞工学センター所属
1998年~ ドイツ生物医学工学会細胞組織工学WG議長
2001年~ 国際医学・生物工学連盟細胞工学WG議長
2003年~ ドイツ教育研究省指名科学評価委員
2003年~ 生物工学強化センター議長
[受賞]
1989年 ドイツ生物医学工学会表彰
1991年 上海生物物理学会名誉会員
アーヘン応用科学大学の紹介
1971 年設立。8,500人を越える学生と200人の教授が2地域(アーヘンとユーリッヒ)に在籍。都市工学・航空工学・IT技術・デザインや経営工学などの工学分野をはじめ、生体医工学・細胞分子生物学などの最先端のバイオ研究などが様々な大学と国際的に連携しながら行われている。
CT&T(細胞組織技術研究所)の紹介
アートマン教授により2001年創設。生物工学・細胞工学・生物物理学・分子生物学などの最先端技術を用いて医薬・医療・バイオ関連産業などへの応用・実用化を目指した開発が、様々な先端研究機関と国際的に連携しながら行われている。
参考資料
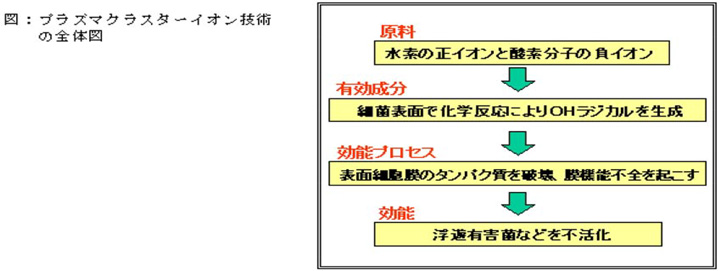
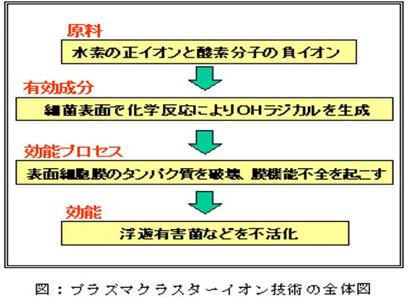
プラズマクラスターイオン技術の全体像
プラズマクラスターイオン技術の有効成分、効能プロセスを実証したことにより、技術の全体像(原料、有効成分、効能プロセス、効能)が明確になりました。
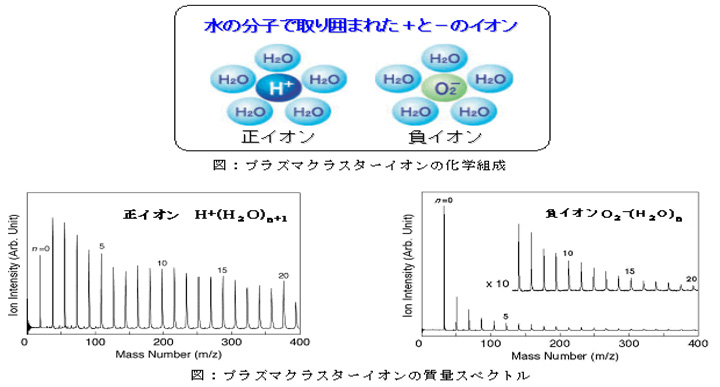
【原 料】プラズマクラスターイオンのイオン種確認
プラズマクラスターイオン発生素子を室内環境下で動作させ、素子より生成されたイオンの質量数を飛行時間型質量分析法で観測しました。その結果、正イオンは水素イオンでその周りに水分子が付いているクラスターイオン、負イオンは酸素分子イオンでその周りに水分子が付いているクラスターイオンであることを確認。また、これらのイオン種は自然界に存在しているイオンであることが知られています。下図はプラズマクラスターイオンの質量を示したものです。下左図は正イオンの質量数(横軸)を示しています。正イオンは質量数19から37、55、73・・・と18(水の質量数)間隔でピークが現れ、H+(水素イオン)に水が付いています。下右図は負イオンの質量数(横軸)を示しています。負イオンは質量数32から50、68、86・・・と18(水の質量数)間隔でピークが現れ、O2−(酸素分子イオン)に水が付いています。
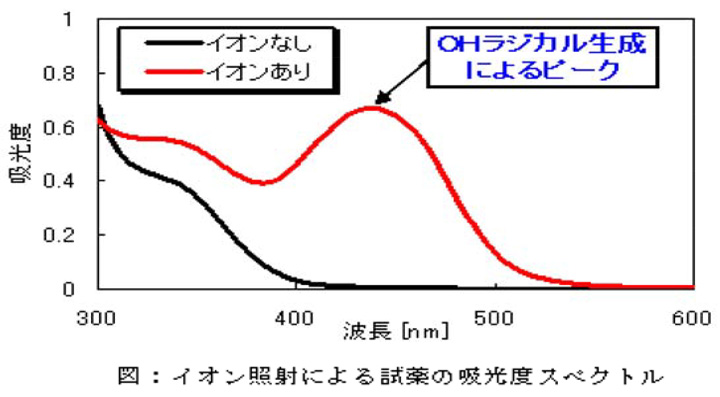
【有効成分】イオン反応による活性種の生成確認
容積4Lのボックス内にプラズマクラスターイオン発生素子と試薬を入れたシャーレを一定時間ボックス内に設置。イオンを照射後、試薬を分光分析しました。その結果、イオンを照射することによりイオン反応で生成されたOHラジカルと試薬が反応し、OHラジカル生成によるピークが現れることを確認しました。
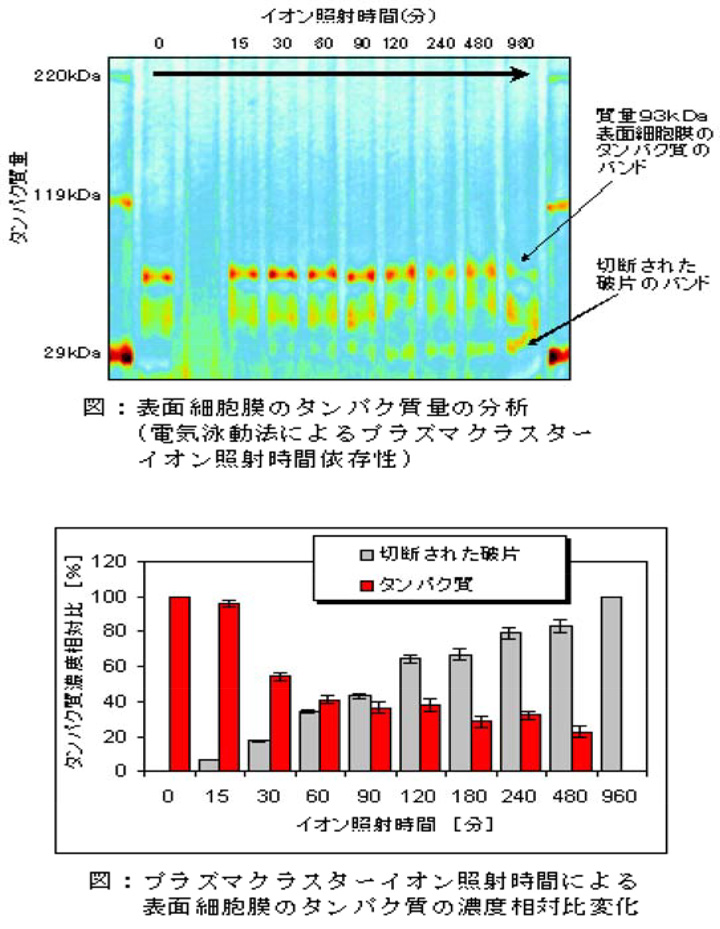
【効能プロセス】細菌の表面細胞膜のタンパク質のフラグメント化(切断)効果
容積4Lのボックス内にプラズマクラスターイオン発生素子を設置し、細菌に一定時間イオンを照射後、細菌の表面細胞膜のタンパク質を分析しました。その結果、イオン照射時間変化により質量93kDa表面細胞膜のタンパク質部分のバンドが減少し、質量34kDa部分にバンドが現れました。これはイオンが93kDa表面細胞膜のタンパク質を34kDaに切断していることを意味します。これにより細菌の不活化は表面細胞膜のタンパク質が破壊されて膜機能不全が起こることによるものと確認しました。
※こちらのページに記載されている内容は、報道発表日時点の情報です。ご覧になった時点で、内容が変更になっている可能性がありますので、あらかじめご了承下さい。